
Research content
研究内容

ヒトは年を経るにつれ確実に老化しますが、周りを見渡せば年齢の割に若く見られる方、 運動能力を維持されている方、比較的年配に見られる方など様々です。その違いを もたらす原因の一つとして食習慣の違いがあげられます。つまり、日々摂取する各種 栄養素は、極端に多すぎても、逆に少なすぎても健康を維持することは難しく、生活 習慣病と呼ばれる病気の原因となります。私共は糖尿病・肥満・メタボリックシンドローム と言った生活習慣病および老化に伴って発症する病気のメカニズムを調べ、食品の 各種機能性を利用して予防する研究を行っています。




メイラード反応
還元糖と蛋白の非酵素的な縮合反応は、一般にグリケーション(glycation)、もしくは発見者の名にちなんでメイラード反応と呼ばれており、大略するとアマドリ転位産物を生成する前期反応と、その後Advanced Glycation End products (AGE)へと変化する後期反応に分けられます。当初メイラード反応は、食品の加工、貯蔵する際に起こる褐変反応として広く研究されてきましたが、ここ十数年間の研究によって、AGEが種々の生体組織に加齢に依存して蓄積すること、また、これらAGE蓄積が糖尿病合併症や動脈硬化などの加齢関連疾患で有意に増加すること、さらに生体はこれらAGE蓄積に対して受容体を介する排除・処理機構を有していることが明らかになってきました。
現在提唱されている主な老化仮説として
(1) プログラム説
(2) エラー破局説
(3) 蛋白の翻訳後修飾説
などが挙げられていますが、メイラード反応は蛋白の翻訳後修飾に重要な役割を果たしていると考えられております。2001年にヒトの全ゲノムが解読され、今はポストゲノム時代と言われているように、mRNAから蛋白への翻訳及びその翻訳後修飾に脚光が浴びるようになってきました。当研究室では、生体における主要AGE構造体の同定及び、AGEと病態との関与、AGE生成メカニズム、AGE受容体の意義を解析し、今後、増々増加すると予想される加齢を基盤とする慢性疾患の予防および治療に貢献できることを目標にしております。
食品成分を用いた動脈硬化予防
天然物抽出エキスおよび天然物由来精製化合物からヒト単球由来マクロファージの泡沫化を阻害する物質をスクリーニングした結果、トマト果実より発見されたステロイドアルカロイド配糖体であるesculeosideA(エスクレオサイドA)のアグリコン(糖が外れた構造)であるesculeogeninA(エスクレオゲニンA)が、マクロファージの泡沫化を顕著に抑制することが明らかとなった。




トマト由来の新規動脈硬化抑制化合物
血管内膜下にプラークが発生して血流が悪くなる状態である。最終的に、動脈の血流が遮断され、酸素や栄養が組織に到達できなくなる結果、脳梗塞や心筋梗塞の原因となる。
初期の動脈硬化病変では、細胞内に多量のコレステロールエステルを蓄積したマクロファージ由来泡沫細胞の集積がみられる。この泡沫細胞の形成が動脈硬化の発症に重要な役割を果たしていることが知られている。
ゆえに、このマクロファージの泡沫化(泡沫細胞の形成)を抑制することによって、動脈硬化の予防が可能になると考えられている。
天然物抽出エキスおよび天然物由来精製化合物からヒト単球由来マクロファージの泡沫化を阻害する物質をスクリーニングした結果、トマト果実より発見されたステロイドアルカロイド配糖体であるesculeosideA(エスクレオサイドA)のアグリコン(糖が外れた構造)であるesculeogeninA(エスクレオゲニンA)が、マクロファージの泡沫化を顕著に抑制することが明らかとなった。


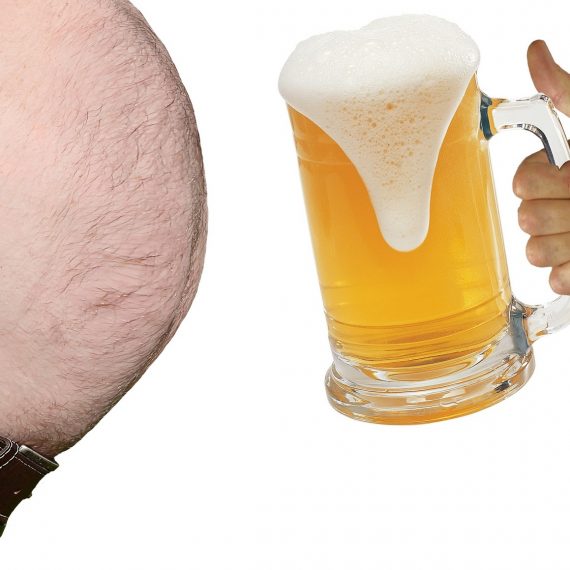
食品成分を用いた脂肪細胞機能改善
脂肪細胞の機能障害は、動脈硬化症および糖尿病合併症の発症に大きく関与している“要”であることがわかる。そこで、脂肪細胞の機能改善を行うことで、メタボリックシンドロームの発症が抑える可能性が考えられる。


コレステロール代謝
ACATはコレステロールの3位のアシルCoAから脂肪酸を転移し、コレステロールエステル合成を触媒する細胞内小胞体酵素です。 一般に生体内では、4つの臓器で重要な生理的役割を持っています。肝臓においてはACATにより合成されたコレステロールエステルはVLDL(超低比重リポタンパク質)の中心に存在する構成脂質として組み込まれます(アセンブリー)。小腸においてはコレステロールが小腸上皮に吸収され、ACATによってコレステロールエステルに変換されカイロミクロンの構成脂質として組み込まれ肝臓へ搬送されます。
○トマト新規配糖体esculeosideAとトマト新規サポゲノールesculeogeninA
ミニトマト果実より単離されたesculeosideAの含有量は1kgあたり400~500mgで、トマトを代表する成分であるlycopene(1kgあたり60mg)よりも多い。さらに、日本で売られているトマト(桃太郎トマト、ミディトマト等)のほぼ全てに含まれていることが明らかになっている。
また、esculeosideAを酸加水分解することで、その糖鎖を切断し、esculeosideAのアグリコンである新規サポゲノールesculeogeninAが得られている。一般的に天然物に含まれる配糖体は、経口投与された後、腸内細菌の働きで糖鎖部分が切断され、生理活性物質としてアグリコンが作られて吸収される。つまり、esculeosideA は、通常の配糖体と同様にesculeogeninAに変換された後に吸収され、esculeogeninAが生理活性物質として働くと考えられている。

○EsculeogeninAのコレステロールエステル(CE)蓄積に対するメカニズム
動脈硬化の初期病変であるマクロファージの泡沫化(細胞内CEの蓄積)を抑制する天然物由来化合物の探索がなされ、esculeogeninAが強い抗泡沫化作用を示した。
CE蓄積に対する抑制メカニズムの解析が行われた結果、esculeogeninAはアセチルLDLの細胞内への取り込みには影響を及ぼさなかったが、ACAT-1およびACAT-2の酵素活性を顕著に抑制した。ACATはコレステロールの細胞内貯蔵やリポタンパクアセンブリー、ホルモン合成、コレステロール吸収において重要な役割を果たしている。つまり、esculeogeninAはACATの活性を抑制することで、マクロファージの泡沫化を抑制することが示唆された。
高脂血症モデルマウスであるapoE欠損マウスを用いた生体における効果の検討から、esculeosideAを90日間経口投与すると、体重の変化は認められないが、総コレステロール値およびLDLコレステロール値、トリグリセリド値が有意に低下が確認された。また、毒性試験からはこの抑制作用は細胞毒性によるものではないことが確認されている。さらに、esculeosideAは大動脈起始部における動脈硬化部位の面積を有意に減少させた。
これらの結果から、esculeosideAは体内でesculeogeninAに変化し、動脈硬化の進展を予防することが示されている。
Fujiwara Y., KiyotaN., HoriM., MatsushitaS., Iijima Y., Aoki K., Shibata D., TakeyaM., IkedaT., NoharaT and Nagai R. Esculeogenin A, a new tomato sapogenol, ameliorates hyperlipidemia and atherosclerosis in apoE-deficient mice by inhibiting ACAT. Arterioscler Thromb Vasc Biol. 27, 2400-2406 (2007)
現在は、トマトからesculeosideAの単離、HPLCによるesculeosideA の分析、esculeosideA高含量トマトの探索を行っております。

ACATとは
ACATはコレステロールの3位のアシルCoAから脂肪酸を転移し、コレステロールエステル合成を触媒する細胞内小胞体酵素です。
一般に生体内では、4つの臓器で重要な生理的役割を持っています。肝臓においてはACATにより合成されたコレステロールエステルはVLDL(超低比重リポタンパク質)の中心に存在する構成脂質として組み込まれます(アセンブリー)。小腸においてはコレステロールが小腸上皮に吸収され、ACATによってコレステロールエステルに変換されカイロミクロンの構成脂質として組み込まれ肝臓へ搬送されます。ステロイドホルモン産生細胞ではステロイドホルモンの原料としてコレステロールをコレステロールエステルとして貯蔵しています。動脈硬化病変ではマクロファージの泡沫化に深く関与しています。
組織分布
ACAT-1とACAT-2の生体内における発現については動物種による違いが見られます。ACAT-1 は、ヒトを含む霊長類、マウス、ラットにおいて生体内に広く分布し、特に副腎、マクロファージなどに高発現しています。一方、ACAT-2 はヒトにおいては主に小腸(と胎児肝臓)に、ヒトを除く霊長類、マウス、ラットなどでは小腸および肝臓に限局しています。
ACATのクローニング
1993 年、約10年の歳月をかけてDartmouth大学のTY ChangらによりヒトACATのcDNAのクローニングに成功してから、ACATに関する研究がこの10年間で飛躍的に発展しました。その後ノックアウ トマウスの解析によりACATにはアイソザイムが存在する事が示唆され、TYChangらによりクローニングされたACATはACAT-1と命名されてい ます。その後マウス、ラビット、ハムスター、モンキー、ラットのACAT-1がクローニングされている。またノックアウトマウスの解析によりアイソザイム の存在が示唆されたため、クローニング競争が展開され、1998年にヒト、マウス、モンキーのACAT-2のシークエンスが同時に発表されました。我々の 教室ではラットACAT-1のクローニングに引き続き(1998年)、昨年ラットACAT-2のクローニングに成功ております。
Hydrophobicity 解析によりACATは膜貫通蛋白と推定され、ヒトACAT-1の膜貫通領域は少なくとも7カ所と想定されました。実際にchangのグループは hamagglutinin(HA)を用いた解析で膜の配向を決定しています。またRudel等はモンキーのACATでは、ACAT-1、ACAT-2共 に膜貫通領域は5箇所と推定し、ACAT-1では269番目の、ACAT-2では249番目のセリンを変異させることにより、ACAT活性が消失することを見いだしました。興味深いことに活性中心思われるセリンはACAT-1では細胞質内に面し、ACAT-2は小胞体ルーメンに面していました。この解析よ りアイソザイム間での機能の違いが活性中心の位置の違いにより生じていることが想定されました。

ACAT-1の調節
ヒト動脈硬化病変ではACAT-1の高度の発現が認められ、細胞特異的モノクローナル抗体を用いた二重染色により、ヒト動脈硬化病変におけるACAT-1発現細胞は、マクロファージが主であることを我々は観察しました。
またヒト単球からマクロファージへの分化に伴い ACAT-1 mRNA および蛋白質発現が5~10倍にも上昇することを我々と他のグループが報告しています。以上のことから、単球・マクロファージにおけるACAT-1発現調節の解析が進んでいます。これまでに単球・マクロファージにおいて複数の因子が ACAT-1 蛋白の発現を誘導することが報告されています。ACAT-1蛋白の発現促進はmRNAレベルであることが我々のグループも含めて数多くの報告があります。
単球・マクロファージの分化に伴うACAT-1 発現上昇や分化過程での複数の因子による ACAT-1発現誘導においては、4つの mRNA のうち 2.8 kb と 3.6 kb の mRNA の選択的な発現上昇を伴うことが多く報告されています。ACAT-1 mRNA は P1 と P7 の2つのプロモーターによりその発現調節が行われていることが知られています。そのうち P1 プロモーターは第1染色体に、一方 P7 プロモーターは第7染色体に存在します。 ACAT-1 mRNA のうち2.8 kb、3.6 kb の2つの mRNA は、P1 プロモーターによる調節を受けており、4.3 kb mRNA は P1 と P7 の2つのプロモーターにより調節を受ける trans-splicing という新たな mRNA の組み換え機構によって生成すると考えられています。7.0 kb の mRNA の発現調節機構についてはいまだわかっておりません。このことから、少なくとも単球・ マクロファージの分化過程における ACAT-1 mRNA の発現上昇には、 P1 プロモーターが深く関わっていることが示唆されます。しかしながらアディポネクチンは ACAT-1 の4つすべての mRNA の発現を抑制することを見いだしました。このことから、アディポネクチンによるヒト単球由来マクロファージの ACAT-1mRNA 発現抑制には、これまで報告されている ACAT-1 mRNA 発現上昇とは異なる機序の関与が考えられます。
ACAT 阻害剤
ACAT は動脈硬化病巣の形成、食事性コレステロールの吸収、リポ蛋白合成などにおいて重要な働きを担っています。そのため、ACAT の活性を阻害する ACAT 阻害剤は、血中コレステロール低下作用、泡沫細胞形成抑制を介した動脈硬化治療薬として期待されています。これまでに多くの ACAT 阻害剤が開発されています。
ACAT 阻害剤は、FR145237 , F-1394, Dup128, E5324, CI277082, NTE-122 などのようにその構造中にウレア基(H2N-CO-NH2) を有するウレア剤と、58-035, CI-976, CI-1011 などアミド基(-NH-CO-)を有するアミド剤の2つに大別されます。
ACAT 阻害剤の一般的な特徴としては、環状構造とアルキル鎖を持つものが多く、疎水性が高いです。ACAT の基質は、環状構造を持つコレステロールと脂肪酸が活性化されたアシルコエンザイム A でありますが、それらの物理化学的性質はACAT 阻害剤と類似しているとみなすこともできます。ACAT の役割は、過剰の細胞内遊離コレステロールをエステル化し、細胞内コレステロールレベルを一定に保つことでありますが、臓器により異なる役割を担っています。このため ACAT 阻害剤には臓器特異的な効果が期待されます。ACAT 阻害剤は、コレステロールエステルの過剰蓄積を阻害し、マクロファージの泡沫化抑制、さらには動脈硬化の発症・進展を抑えることが期待されます。これまでに動物や初代培養系のヒト単球由来マクロファージを用いた検討から、CI-1011、58-035などのACAT 阻害剤がマクロファージの泡沫化抑制作用や抗動脈硬化作用を有することが報告されています。
スルホニルウレア剤(SU 剤)は、膵臓β細胞のスルホニルウレア受容体(SUR)への結合によるインスリン分泌を促進させる2型糖尿病治療薬です。SU剤が膵臓β細胞以外の細胞で有用な薬理効果を発揮することが最近報告されています。例えば、肝臓における糖新生の抑制、末梢組織におけるインスリン感受性の増強などが知られています。また、胆肝上皮細胞における胆汁分泌促進作用やサイトカイン誘導性の好酸球活性化に対する抑制作用、抗癌剤耐性に関する P-糖タンパク質の機能阻害、血小板凝集抑制作用、抗酸化作用に基づく抗動脈硬化作用などが報告されています。このような背景をふまえ、我々は SU 剤の膵外作用に注目し、J774 細胞(マウスマクロファージ細胞株)、ACAT 過剰発現細胞を用いた検討でSU 剤であるグリベンクラミドが ACAT 阻害剤として作用することを明らかにしました。
肝臓では、ACAT によって合成された CE をはじめ、トリグリセリド、リン脂質などVLDL を構成する脂質は小胞体で合成されます。これらの脂質はミクロソームトリグリセリド転送タンパク(microsomal triglyceride transfer protein : MTP)等の働きにより VLDL に組み込まれ、血中に放出されます。ACAT 阻害剤により肝臓のコレステロールエステル合成を阻害しますと、VLDL の肝細胞内分解が促進され、細胞外への VLDL 分泌が抑制されることが報告されています。小腸では、腸管からコレステロールが小腸上皮に吸収され、ACAT によって CE に変換された後、カイロミクロンの構成脂質として組み込まれます。ACAT 阻害剤により小腸上皮におけるコレステロールのエステル化が抑制されると、コレステロールの吸収が阻害されます。したがって、ACAT 阻害剤が小腸で働くと、コレステロール吸収阻害による血中コレステロール低下作用が期待されます。
ACAT欠損マウスモデル
1996 年にACAT-1欠損マウスが報告され、確かにマクロファージや副腎のACAT活性は消失し、肝臓や小腸のACAT活性は野生型と変わりませんでした。このマウスの特徴的な所見として副腎と培養マクロファージのCEの減少が観察されました。2000年に報告されたACAT-1欠損マウスはマイボーム腺の萎縮と眼瞼の狭小化が観察されました。さらにアポE又はLDL受容体欠損マウスとの交配させた動脈硬化モデルマウスが二つのグループから報告されました。これらのダブル欠損マウスでは全身の皮膚黄色種や皮膚の肥厚、脱毛を呈しました。動脈硬化病変の検討では有意に減少したと報告するグループに対して、もう一方のグループでは病変部の構成が変化したことのみ報告しています。さらにFazioらはマクロファージに特異的なACAT-1欠損が動脈硬化に及ぼす影響を検討しました。驚くことにACAT-1欠損マクロファージの移植を受けた群では動脈硬化が促進されるというものでありました。
次に2000 年にACAT-2欠損マウスが報告されました。このマウスでは高脂肪食負荷による高脂血症に耐性を示し、コレステロール胆石の形成も抑制されました。高脂肪食下ではコレステロールの吸収が著明に抑制され、ACAT-2は食事由来のコレステロール吸収の制御に重要な役割を演じていることが示唆されました。 2003年にはアポE欠損マウスとの交配による動脈硬化モデルマウスが作成され、ダブル欠損マウスでは動脈硬化病変が有意に抑制されました。これらの知見からACAT-2の選択的な阻害剤が食事性高脂血症の治療としてなりうる可能性が示唆されました。
メタボリックシンドローム予防を目的とした脂肪細胞の機能改善に関する研究
≪メタボリックシンドロームの発症メカニズム≫
*図の概要*
肥満状態では脂肪細胞が肥大化し、非酵素的翻訳後修飾も進行して(Nagai R et al., J. Biol. Chem. 2007)、脂肪細胞の機能障害が起こる。その結果、善玉サイトカインであるアディポネクチンの分泌低下からインスリン抵抗性が惹起され、糖尿病、高脂血症、高血圧の発症頻度が上昇する。これらは単独でも動脈硬化症の原因になるが、併発すると動脈硬化症の発症率は格段に上昇する。
つまり、脂肪細胞の機能障害は、動脈硬化症および糖尿病合併症の発症に大きく関与している“要”であることがわかる。そこで、脂肪細胞の機能改善を行うことで、メタボリックシンドロームの発症が抑える可能性が考えられる。
≪内臓脂肪の肥大化と脂肪細胞の機能低下の関連≫
不規則な生活をしていると・・・
運動不足・過栄養・遺伝的素因 ⇒ 内臓脂肪の肥大化
アディポネクチン:脂肪細胞から分泌される善玉サイトカイン。インスリン感受性を高めたり、脂肪を燃焼させたり、動脈硬化の予防作用があると考えられている。 ※アディポサイト=脂肪細胞
肥大化した脂肪細胞ではアディポネクチンの分泌量が低下する。
また、脂肪細胞がフマル酸によって翻訳後修飾を受けることによって(Nagai R et al., J. Biol. Chem. 2007)、脂肪細胞の機能が障害を受けていると考えられている。
≪蛋白のAGE化と類似する、蛋白の2SC化≫
AGE生成メカニズム
遺伝子(DNA)
↓ 転写
mRNA
↓ 翻訳
A蛋白
↓
翻訳後修飾
↓
修飾を受けた蛋白A
↓ ↓
機能調節 機能低下→病態進展
翻訳後修飾には酵素的反応、非酵素的反応の2種類が存在している。
・酵素的反応:秩序のある翻訳後修飾で、リン酸化、アセチル化、糖鎖付加があげられる
・非酵素的反応:無秩序な翻訳後修飾で、脂質過酸化、メイラード反応によるAGE化があげられる。
⇒このように、AGEは蛋白の非酵素的な糖化反応から生成する。
●AGEと類似する2SCとは?
蛋白を構成するアミノ酸の一種「Cystein(システイン)」と「フマル酸」の翻訳後修飾反応(ミカエル付加反応)から生成される物質である。
Cystein + フマル酸 → S-(2-succinyl)cysteine (2SC)
(タンパク質)
≪アディポネクチンの2SC化について≫
脂肪細胞が分泌している善玉サイトカインであるアディポネクチンにもCysteinが存在している。また、高濃度グルコース下では脂肪細胞内でフマル酸含量が顕著に増加し、アディポネクチンが2SC化する。
≪研究目的≫
●2SC化抑制化合物の探索
食品から以下の効果があるものを探索する。
①細胞内フマル酸濃度を下げる
②アディポネクチンの2SC化を阻害する
天然素材である食品から見つけだすことで、安全にメタボリックシンドロームの進展が予防できればと考えている。
【研究報告】
現在4年生が習得したこと
・3T3-L1(繊維芽細胞)の継代培養の手法
・3T3-L1の分化による脂肪細胞(adipocyte)の作製手法。
・脂肪細胞染色
・細胞保存法
・2SCの合成
・ELISAによる2SC化蛋白の検出
・アミノ酸分析